Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Klebsiella pneumoniae (K. pneumoniae) pose an emerging threat to public health sector worldwide. They are one of the potent nosocomial pathogens and cause variety of infections including pneumonia, septicaemia, wound infections, urinary tract infections and catheter-associated infections. From the last two decades, these pathogens are becoming more powerful due to the acquisition of resistomes on different types of plasmids and transposons. There are four main mechanisms of antibacterial resistance such as efflux pump, target alteration, membrane permeability and notably enzymes hydrolysis. K. pneumoniae produce different types of enzymes but most importantly extended spectrum-β-lactamase (ESBL), carbapenemase and metallo-β-lactamase (MBL). K. pneumoniae carbapenemases (KPCs) and New Delhi metallo-β-lactamase (NDM) producing isolates displayed resistance not only against the β-lactam drugs (penicillins, cephalosporins and carbapenems) but also to other classes of antibiotics (aminoglycosides and quinolones). Therapeutic options available to treat serious infections caused by these extensively drug-resistant pathogens are limited to colistin, tigecycline and fosfomycin. Hence, combination therapy has also been recommended to treat such bacteria with clinical side effects, therefore, new treatment regime must be required. Moreover, we are relying on conventional diagnostic tools, however, novel techniques must be required for robust identification of multi-drug-resistant bacteria.
Department of Microbiology, Faculty of Life Sciences, Government College University, Pakistan
Muhammad Hidayat Rasool
Department of Microbiology, Faculty of Life Sciences, Government College University, Pakistan
Shah Jahan
Department of Immunology, University of Health Sciences, Pakistan
Muhammad Shafique
Department of Microbiology, Faculty of Life Sciences, Government College University, Pakistan
Bilal Aslam
Department of Microbiology, Faculty of Life Sciences, Government College University, Pakistan
*Address all correspondence to: musmanqamar@gcuf.edu.pk
1. Introduction
Klebsiella pneumoniae is one of the notorious pathogens with increase number of severe infections and shortage of effective treatment regimes. They are extremely resilient pathogens because their success seems to follow the model of “the best defence for a pathogen is a good defence” rather than “the best defence for a pathogen is a good offense”. K. pneumoniae is a Gram negative, non-motile, capsulated and an imperative member of Enterobacteriaceae family. They were firstly identified in late nineteenth century and given the name Friedlander’s bacterium [1]. They are hermaphrodite and cause both community acquired infections (soil, surface water) and significantly hospital acquire infections (medical devices, catheters). Importantly, they can reside on different body mucosal surfaces of oropharynx and gastrointestinal tract and cause variety of infections including pneumonia, bacteraemia, septicaemia, gastritis, wound infections, urinary tract infections, and soft tissue surgical infections [2]. From the last 2 decades, K. pneumoniae strains acquired extra genetic traits and become more virulent and antibiotic resistant [3]. They develop resistance against a wide range of antibiotics including carbapenems. Therefore, they produce (or synthesise) extended spectrum-β-lactamases (ESBL), carbapenemases, metallo-β-lactamases (MBLs), New Delhi metallo-β-lactamases (NDM) and have many other resistant mechanisms such as efflux pump and membranous target alteration.
Antimicrobial resistance (AMR) defined when a bacterial strain shows resistance against antibiotics that normally inhibit or stop their growth and allow them to withstand against drugs. Last 2 decades, there are various AMR pathogens have been emerged around the globe such an ESBL producing pathogens, multi-drug resistant and extensively drug resistant Mycobacterium tuberculosis (MDR, XDR-MTB), MBL producing bacteria, methicillin resistance Staphylococcus aureus (MRSA) and the advent of NDM producing bacteria. These isolates are almost impossible to treat with the existent treatment regime [4]. AMR emerged even since the discovery of first antibiotic “penicillin” in 1940 due the natural process of bacteria. Certainly, the genes that confer resistance upon some strains of bacteria precede antibiotics by millions of years [5]. Currently, AMR is becoming a big health issue around the world due to the irrational, overused and misuse of antibiotics which can lead to extra financial burden, prolong hospital stay and even fatal consequences. Unfortunately, currently we deficiency of the novel effective antimicrobial compounds, peptides and pipelines antibiotics to combat with emerging superbugs which result us facing growing enemy with largely depleted armoury [6].
3. Mortality, morbidity and economic burden due to antimicrobial resistant bacteria
AMR pathogens are one of the main cause of morbidity and mortality in both developed and developing countries. According to a published data, around 700,000 people die due to infections caused by AMR pathogens every year worldwide and most of the data is still underreported because of poor reporting and surveillance studies [7]. On the other hand, as per The Centre of Disease and Control (CDC), above 2 million people get infected due to AMR bacteria with a mortality rate of 23,000 each year [8]. Similarly, an Indian study revealed that around 60,000 neonates die due to AMR infections per year [9]. Likewise, a study conducted in Pakistan in 2009 was also documented that 40% neonates die due to MDR and XDR pathogens (Escherichia coli, K. pneumoniae, Enterobacter cloacae) isolated from blood samples [10]. Another study from Pakistan testified that 37 out of 78 new-borns died due to infections caused by MDR-Acinetobacter spp., [11]. Qamar et al., conducted a study in Islamabad, Pakistan also reported 4/9 children died due to NDM positive isolates mainly of E. coli and K. pneumoniae [12]. Recently, Khan et al., from Karachi, Pakistan also reported that 22 with <1 month of age and 10 were elderly patients died due to bacteraemia and septicaemia which was mainly caused by NDM positive K. pneumoniae [13]. Highest mortality rate is mainly reported in developing countries as compare to developed countries largely in immunocompromised persons. Major risk factors in public sectors hospitals of developing countries are, poor hygienic practices of health care workers, irrational use of antibiotics, poor sanitation and hygienic facilities, nosocomial pathogens, hospital and wards are overcrowded with patients and attended and lack of health care workers, floors and rooms are spilled with sputum and operation theatres are in deliberated condition and are poorly equipped, contaminated drinking water, more than one patients on single bed, presence of insects and flies and most importantly low health care budget by the Government [14].
Every year, millions of dollars are spent to tackle the burden of AMR bacterial infections around the globe. European Union invested around €1.5 billion annually to tackle the infections caused by AMR pathogens which are responsible for around 23,000 mortalities only in Europe [15]. However, United States of America also expenditures above $20 billion to treat the 20 million patients who get infected with AMR infection each year [16].
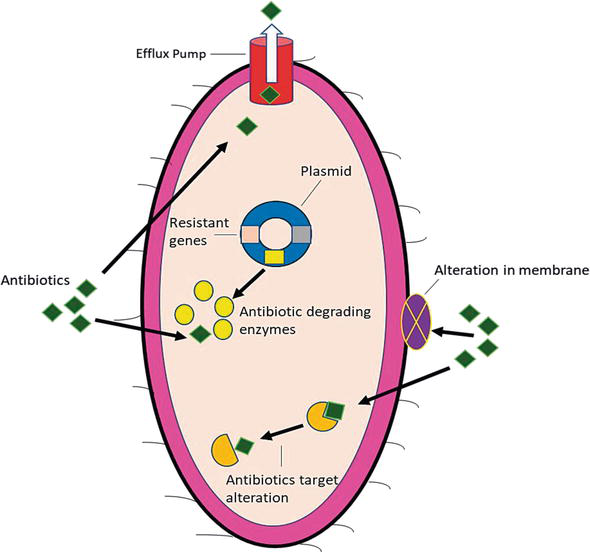
Since 1990s, bacteria have been becoming gradually more lethal, due to acquisition of resistomes and develop resistance to different sets of antibiotics. Now a day, many bacterial pathogens notably K. pneumoniae are intrinsically resistant to various antibiotics or they acquire or encoded antimicrobial resistant genes mainly on plasmids as well on chromosomes [17]. There are different mechanisms such as conjugation, transformation and transduction in which susceptible strains can acquire resistance genes with transposons that eventually support different resistance genes to incorporate with host chromosome or plasmids. Presently, there are four fundamental mechanisms of bacterial drug resistance (1) target alteration (2) modification in membrane permeability (3) efflux pump and (4) antibiotic degradation by enzymes as shows in Figure 1 [18]. In K. pneumoniae both the efflux pump and enzymatic degradation of different antibiotics are more efficient as compare to other two mechanisms.
Figure 1.
Mechanism of antimicrobial resistance.
4.1 Target alteration of the antibiotics
In general, many antibiotics bind to different bacterial target sites with high affinity. So, most of the bacteria including K. pneumoniae alter the target sites and structure to block or inhibit the efficient binding sites [19]. In K. pneumoniae, antibiotic resistance occurs due to target modification in lipopolysaccharides (LPS), which is interceded by the activation of PmrA/PmrB two component regulatory system (TCRS). Moreover, another TCRS, PhoP/PhoQ contributes to produce polymyxin resistance by indirectly activating the PmrA/PmrB TCRS via PmrD linker protein. Similarly, inactivation of the PhoQ/PhoP negative regulator encoded by mgrB gene has also been identified to play a prominent role in polymyxin resistance in K. pneumoniae [20]. There are various pathogens that cause different infections and develop resistance even against single point mutation of resistant genes that encode an antibiotic target. These resistant strains then further produce numerous copies of bacteria [21].
4.2 Modification in membrane permeability
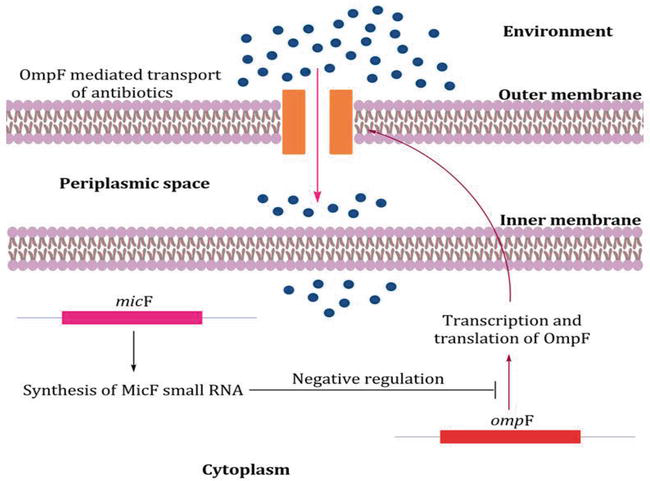
Outer membrane of the bacteria is first line barrier to protect the microorganisms against unfavourable environment including chemical and biological materials. In comparison, majority of the Gram-negative rods (GNR) are intrinsically less permeable to various antibiotics because of outer membrane barrier than Gram positive bacteria [22]. In the cell wall of Enterobacteriaceae family including K. pneumoniae contains various outer membrane proteins (Omps) such as OmpA, OmpF and OmpC and the hydrophilic antibiotics can cross through these Omps by diffusion. These proteins are involved in adhesion to other cell surfaces, maintenance of cell structure and regulation of transport of bactericidal agents and nutrition. OmpF expression is regulated at the posttranscriptional level via the micF RNA, and many different environmental stresses impact the OmpC/OmpF ratio via changes in micF levels. The expression of micF is increased along with that of OmpF at high OmpR-P concentrations. The expression of micF is also induced by toxic agents, such as paraquat and weak acids, by the SoxS, MarA, and Rob transcription factors, which bind to the same sequence element in the micF promoter and they down regulate the synthesis of OmpF at transcriptional level (Figure 2) [23]. Therefore, the bacteria showed resistance by limiting the antibiotics entry in to the cell by the down regulation or replacement of porins with more selective channels. However, recent data showed that the loss of porins expression play a significant role to produce resistance to new drugs (cephalosporins and carbapenems), to which resistance is manly mediated by the enzyme degradation [24]. OmpA is one of the major protein of K. pneumoniae that prevent the activation of airway epithelial and loss of this protein make the bacteria susceptible. There are two important Omps in K. pneumoniae; OmpK35 and OmpK36 that allow the hydrophilic molecules such as nutrients and carbapenems to diffuse into bacteria. Moreover, K. pneumoniae also contain two alternative proteins; KpnO and OmpK26. Any mutation in OmpK36, OmpK26 and KpnO leads to increased resistance to cephalosporin and carbapenems [25].
Figure 2.
Down regulation mechanism of outer membrane proteins (OMPs) in gram negative bacteria.
4.3 Increased efflux pumps
Efflux pumps are the active transport mechanisms that expel out antibiotics from bacterial cell and play a vital role to develop intrinsic resistance against broad range of antibiotics in GNR [18]. Currently, there are several MDR efflux pumps are present in bacteria including FuaABC in Stenotrophomonas maltophilia, MedeA in Streptococcus mutans and AcrAB, KexD and more recently OqxAB efflux pumps in K. pneumoniae [26, 27]. In K. pneumoniae, most efficient antimicrobial resistant efflux pump is AcrAB, which is not only expel out the β-lactam, quinolones and chloramphenicol but also the host derived antimicrobial agents such as human antimicrobial peptides and human bronchoalveolar lavage fluid [25]. Recently, newly emerging OqxAB efflux pump has been recognised in different parts of the world that conferring resistance to quinolones and chloramphenicol [27].
4.4 Enzymatic hydrolysis of the antibiotics
In K. pneumoniae, the most potent and significant antimicrobial resistant mechanism is enzymatic hydrolysis of wide range of antibiotics including β-lactam, quinolones, aminoglycosides and macrolides [18]. β-lactamases are the enzymes that hydrolyze the effect of β-lactam drugs by attacking on their β-lactam ring. Currently, a total of 4037 β-lactam enzymes have been identified and mong these 1327 are class A, class B (n = 581), class C (n = 1359) and 660 belong to class D [28]. K. pneumoniae encode for many β-lactamases such as ESBLs, carbapenemases and MBLs. ESBLs and carbapenemases contain important subclasses of β-lactamase enzymes such as imipenemase (IMP), Verona integron encoded metallo-β-lactamase (VIM), K. pneumoniae carbapenemases (KPC), oxacillinase (OXA) and recently NDM [29].
4.4.1 Classification of β-lactamases
These enzymes are broadly classified into two main scheme one is Ambler Molecular classification scheme which is based on amino acid sequence and is widely acceptable and secondly, Bush Jacoby classification scheme which is based on biochemical properties [30]. Ambler class is further divided in to four major sub-classes A, B, C, and D based on conserved and distinguished amino acid motifs. Among these, class A, C, and D are the serine carbapenemases that hydrolyzed their substrate through active site by forming acyl groups and require serine as a cofactor for their activity. However, class B enzymes are the MBLs that utilised at least one zinc ion at their active site to hydrolyze the β-lactam antibiotics. MBLs are further classified into three super families B1, B2 and B3 [31]. However, Bush-Jacoby-Medeiros classification scheme is divided into three major groups; class C (Group 1 cephalosporinases), class A and D (Group 2 Serine β-lactamases) and class B (Group 3 MBLs). These are further divided in to several different subgroups as shows in Table 1 [30].
Molecular class (subclass)
Bush Jacoby groups
Main substrate
Defining characteristic(s)
Representative
Bacteria
A
2a
Penicillins
Increased hydrolysis of benzylpenicillin
PC1
A
2b
Penicillins and early Cephalosporins
Hydrolysis of the penicillins and cephalosporins
TEM and SHV
Enterobacteriaceae (K. pneumoniae, E. coli, E. cloacae)
A
2be
Extended spectrum-β- lactam drugs, monobactam
Hydrolysis of extended spectrum-β- lactam drugs
TEM, SHV and CTX-M
Enterobacteriaceae (K. pneumoniae, E. coli, E. cloacae)
A
2br
Penicillins
Resistant to β-lactam inhibitor
TEM and SHV
Enterobacteriaceae (K. pneumoniae, E. coli, E. cloacae)
A
2c
Carbenicillin
Hydrolysis of carbenicillin
CARB
Enterobacteriaceae (K. pneumoniae, E. coli, E. cloacae)
A
2ce
Carbenicillin and Cefepime
Hydrolysis of carbenicillin and cefepime
RTG
B (B1)
3a
Carbapenems
Carbapenems but not aztreonam
IMP, VIM and NDM
Enterobacteriaceae (K. pneumoniae, E. coli), non-fermenters (P. aeruginosa, A. baumannii)
B (B2)
3b
Carbapenems
Prefer to hydrolyze carbapenems
CphA and Sfh
C
1
Cephalosporins
More hydrolyze the cephalosporins
AmpC,
C
1e
Cephalosporins
Highly hydrolyzed the ceftazidime
GC1, CMY-37
D
2d
Oxacillin
Hydrolysis of oxacillin
OXA-1, OXA-10
A. baumannii
D
2de
Extended spectrum-β- lactam drugs
Hydrolysis of extended spectrum-β- lactam drugs
OXA-11, OXA-50
A. baumannii
D
2df
Carbapenems
Hydrolysis of oxacillin and carbapenems
OXA-23 and OXA-48
A. baumannii
Table 1.
Classification of the β-lactamases.
4.4.2 Extended spectrum-β-lactamase producing K. pneumoniae
ESBL are the enzymes that hydrolyze the extended spectrum drugs mainly cephalosporins. ESBL producing Enterobacteriaceae mainly the K. pneumoniae and E. coli is becoming one of the serious threat to the public health sectors. According to World Health Organisation (WHO), around 50% of ESBL producing K. pneumoniae becomes the endemic in many parts of the world and it causes 30% resistance rate in the community [32]. In 1965, first β-lactamase enzyme Temoneria (TEM-1) and sulphydryl variable (SHV-1) has been identified. In 1983, first case SHV-2 and TEM-3 has been documented in Germany and France respectively. Similarly, in 1989, USA also reported ESBL producing K. pneumoniae and E. coli. Since then, incidence of these enzymes markedly sporadic from one region to another [33]. ESBL producing K. pneumoniae displayed resistance to clinically effective extended spectrum cephalosporins and monobactam and left limited treatment options. Moreover, ESBL producing K. pneumoniae has become major ESBL carrying bacteria associated with nosocomial infections and outbreak during 1990 to 2000. Later, these pathogens acquired variants of ESBL such as CTX-M, TEM and SHV. There are various types of ESBLs have been identified throughout the world. Up till now, TEM (n = 227) are the most predominant type of ESBLs followed by CTX-M (n = 207), SHV (n = 203), VEB (n = 19) and PER (n = 9) [28]. Majority of these ESBLs have been reported in Enterobacteriaceae notably K. pneumoniae and E. coli. ESBLs producing K. pneumoniae have also become an important hospital acquired pathogen. In China, around 50% of the hospital acquired infections are caused by CTX-M-14 and CTX-M-15 ESBL producing K. pneumoniae [34]. Recently, these ESBL producing K. pneumoniae were also reported from community such as water, soil and veterinary settings [35]. In Asia, SHV is the predominant genotype of ESBL and early SHV-5 and SHV-12 enzymes were documented in Korea and Japan. However, in Europe and south east Asia (India and Pakistan), CTX-M-2, CTXM-3, CTX-M-14 and CTX-M-15 producing Enterobacteriaceae at its rise. However, in Japan, SHV-12 and CTX-M-3 and CTXM-14 have been predominantly reported in K. pneumoniae and E. coli. In Indo-Pakistan region, most of the clinical isolates of K. pneumoniae encoded for TEM, CTX-M and SHV. Two highly populated countries in the world (China and India) represents the largest reservoir of CTX-M in the world [36].
4.4.3 AmpC producing K. pneumoniae
AmpC are the β-lactam enzymes that belong to the ambler class C of β-lactam. These enzymes hydrolyze cephalosporins antibiotics and produce resistance against penicillin, second and third generation cephalosporins and cephamycin. Moreover, these enzymes can also convey resistance to combination of these antibiotics along with β-lactam inhibitors. The versatility of the K. pneumoniae is to encode various β-lactam resistant genes on their transferable plasmids that able them to spread AmpC like resistomes in to other species [30]. Plasmid mediated AmpC gene was firstly identify during 1980s in K. pneumoniae. The most common genes of AmpC families that have been reported in K. pneumoniae are DHA, FOX, CMY and MOX types in different parts of the world. Although they are plasmid mediated however, first case of chromosomal mediated AmpC blaCMY-2 containing K. pneumoniae was reported in 2009. AmpC producing K. pneumoniae along with porin loss and increase efflux pump display increase resistance against β-lactam drugs which can lead to the carbapenem resistance in future [37].
4.4.4 Class A carbapenemases producing K. pneumoniae
Since the sporadic spread of ESBL producing K. pneumoniae in different regions, carbapenems which is considering last resort to treat infections caused by such pathogens have widely used. The extensive usage of carbapenems have led the emergence of plasmid mediated carbapenemase genes such as IMP, VIM, KPC and NDM. Carbapenemases are the potent enzymes that hydrolyzed the carbapenem drugs and left no treatment option available [32]. There are various chromosomal mediated (SME, SFC and NMC-A) and plasmid mediated carbapenemases (KPC, GES, IMI) have been identified. It has determined that a carbapenemase has variants extending from KPC-1 to KPC-33 till now [28]. KPC gene is mainly located on different types of transmissible plasmids such as IncF, IncX, IncA/C, IncR and IncI2 which lead to the horizontal transfer of antimicrobial resistomes. These plasmids also acquired resistant genes of other antibiotics such as aminoglycosides, tetracycline and quinolones. Most of the KPC genes are associated with Tn4401 transposons which jumped from one plasmid to other [38]. the endemic of KPC producing K. pneumoniae identified in different parts of the world including USA, UK, Japan, China, Greece, India, Spain, Germany and Italy. Moreover, out breaks of the KPC also documented in different countries like USA, France (travelling history from the endemic area), Spain, Netherlands, UK, Finland and Sweden [39].
4.4.5 Class B carbapenemase producing K. pneumoniae
Class B are the MBLs that require Zn+ as a cofactor for their activity. There are many types of MBLs have been identified such as IMP, VIM, and NDM in Enterobacteriaceae. These enzymes cause resistance to various antibiotics including carbapenems and left no treatment option [40]. NDM is one the potent enzyme of MBL that newly emerged in clinically isolates of K. pneumoniae and E. coli from a Swedish patient who had been hospitalised in New Delhi, India in 2008. NDM producing bacteria displayed resistance not only against β-lactam drugs but also to other 14 classes of antibiotics. NDM producing bacteria notable K. pneumoniae and E. coli are major cause of morbidity and mortality in developing countries. Qamar et al. reported 4/9 children died due to NDM producing bacteria [12]. Similarly, another study from Karachi, Pakistan also documented 57% mortality in neonates due to NDM producing bacteria mainly K. pneumoniae [13]. Up till now there are 19 (NDM-1 to NDM-19) variants have been identified in different parts of the world [41]. Among these NDM-1, NDM-5 and NDM-7 are most prevalent variants with a hyper variable region. NDM are mainly plasmid mediated as compared to chromosome and located on different size and number of plasmids such as IncF, IncX, IncA/C. NDM producing K. pneumoniae are belong to various sequence types such as ST11, ST147, ST258, ST340, ST15 and ST16. More than 50% of the NDM producing K. pneumoniae have been identified from ST11 and ST147. NDM genes are mainly associated with ISAba125 transposons in Enterobacteriaceae family which hypothesised originated from the Tn125 of A. baumannii [42]. Since the discovery of the NDM in Asia it is widely spread in different parts of the world including USA, UK, Middle East, Europe, Africa and Australia. Main reservoir for NDM are the Asian countries like India, Pakistan and China and the Balkan States [39].
4.4.6 Class D carbapenemases producing K. pneumoniae
These carbapenemase enzymes swiftly hydrolyzed the isoxazolylpenicillins drugs such as oxacillin, cloxacillins and dicloxacillin than benzylpenicillin. There are more than 574 variants have been identified around the globe but few of them have the carbapenemase activity. Therefore, class D are reclassified in to 12 main groups; Oxacillinase (OXA)-23, OXA-24, OXA-48, OXA-51, OXA-58, OXA-143, OXA134, OXA-211, OXA-213, OXA-214, OXA-229, OXA-235 [43]. Among these most potent variants is OXA-48 which was firstly identified in K. pneumoniae in Turkey in 2003. Later, this gene spread in different parts of the world such as Libya, Egypt, India, Spain Switzerland, France, Germany, Argentine, Israel, Saudi Arabia, Japan. The highest percentage of OXA-48 has been reported in France, Spain and Saudi Arabia (78%) followed by 88% in Lebanon and 56% in Pennsylvania [39]. Co-existence of OXA-48 like K. pneumoniae has been reported from different countries like Switzerland (OXA-48/NDM-1), Turkey (OXA-48/NDM-1) and India (OXA-181/VIM-5). OX-48 like genes are mainly present on transferable and conjugative group of plasmids (IncL/M and IncL) [39, 44].
AMR is becoming a menace to global health. According to United Nation (UN), in 2050, if we do not tackle the AMR now, more than 10 million people will die each year and the global income cost would be $100 trillion. So, in 2050, one person will die after every 3 seconds [7]. Firstly, it is need of the hour to give a public awareness through paper and electronic media to stop the irrational use of antibiotics. Secondly, people should improve their hygiene practices which will reduce the 60% antibiotics burden to reduce the diarrhoea. Thirdly, more than 70% world’s antibiotics are being utilised in agriculture and live stocks, so overuse of antibiotics must be avoided in these settings. Fourthly, Global surveillance of AMR should be conducted and maintained throughout the world. Fifthly, there are many countries which do not have the rapid and effective diagnostics facilities of microbiology which allow the physician to prescribe the broad-spectrum empirical therapy. Sixthly, there is an urgent need to develop an alternative method to treat the AMR such as vaccines, phage therapy, probiotics, antibodies and lysins. Seventhly, there are shortages of microbiologists, infectious diseases specialists, infections control specialists, vegetarians, pharmacists and epidemiologists. Eighthly, A global funding must be required to support and encourage the less commercially attractive. Ninthly, the world gives better incentive to pharmaceutical companies to develop new and existing antibiotics. Finally, world leading organisations such as WHO, G20, UN must work together with coordination.
References
1.Merino S, Camprubi S, Alberti S, Benedi VJ, Tomas JM. Mechanisms of Klebsiella pneumoniae resistance to complement-mediated killing. Infection and Immunity. 1992;60:2529-2535
2.Rock C, Thom KA, Masnick M, Johnson JK, Harris AD, Morgan DJ. Frequency of Klebsiella pneumoniae carbapenemase (KPC)-producing and non-KPC-producing Klebsiella Species contamination of healthcare workers and the environment. Infection Control. Hospital Epidemiology. 2016;35:426-429
3.Paczosa MK, Mecsas J. Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiology and Molecular Biology Reviews: MMBR. 2016;80:629-661
4.Ventola CL. The antibiotic resistance crisis: Part 1: Causes and threats. P & T: A Peer-Reviewed Journal for Formulary Management. 2015;40:277-283
5.Read AF, Woods RJ. Antibiotic resistance management. Evolution, Medicine, and Public Health. 2014;2014:147
6.Viswanathan R, Singh AK, Basu S, Chatterjee S, Sardar S, Isaacs D. Multi-drug resistant gram negative bacilli causing early neonatal sepsis in India. Archives of Disease in Childhood Fetal and Neonatal Edition. 2012;97:182-187
7.de Kraker MEA, Stewardson AJ, Harbarth S. Will 10 million people die a year due to antimicrobial resistance by 2050? PLOS Medicine. 2016;13:e1002184
8.Lim C, Takahashi E, Hongsuwan M, Wuthiekanun V, Thamlikitkul V, Hinjoy S, et al. Epidemiology and burden of multidrug-resistant bacterial infection in a developing country. Life. 2016;5:e18082
9.Laxminarayan R, Duse A, Wattal C, Zaidi AK, Wertheim HF, Sumpradit N, et al. Antibiotic resistance-the need for global solutions. The Lancet Infectious Diseases. 2013;13:1057-1098
10.Hannan A, Qamar MU, Usman M, Waheed KAI, Rauf K. Multidrug resistant microorganisms causing neonatal septicaemia: In a tertiary care hospital Lahore, Pakistan. African Journal of Microbiology Research. 2013;7:1896-1902
11.Saleem AF, Ahmed I, Mir F, Ali SR, Zaidi AK. Pan-resistant Acinetobacter infection in neonates in Karachi, Pakistan. Journal of Infection in Developing Countries. 2009;4:30-37
12.Qamar MU, Nahid F, Walsh TR, Kamran R, Zahra R. Prevalence and clinical burden of NDM-1 positive infections in paediatric and neonatal patients in Pakistan. The Paediatric Infectious Disease Journal. 2015;34:452-454
13.Khan E, Irfan S, Sultan BA, Nasir A, Hasan R. Dissemination and spread of New Delhi Metallo-beta-lactamase-1 superbugs in hospital settings. The Journal of the Pakistan Medical Association. 2016;66:999-1004
14.Qamar MU, Hannan A, Arshad MU, Arshad M. Metallo-β-lactamase producing Enterobacter cloacae: An emerging threat in neonates. African Journal of Microbiology Research. 2014;8:525-528
15.Davies SC, Fowler T, Watson J, Livermore DM, Walker D. Annual report of the chief medical officer: Infection and the rise of antimicrobial resistance. Lancet. 2013;381:1606-1609
16.Hampton T. Report reveals scope of us antibiotic resistance threat. Journal of the American Medical Association. 2013;310:1661-1663
17.Tenover FC. Mechanisms of antimicrobial resistance in bacteria. American Journal of Infection Control. 2006;34:3-10; 64-73
19.Billal DS, Feng J, Leprohon P, Légaré D, Ouellette M. Whole genome analysis of linezolid resistance in Streptococcus pneumoniae reveals resistance and compensatory mutations. BMC Genomics. 2011;12:512
20.Haeili M, Javani A, Moradi J, Jafari Z, Feizabadi MM, Babaei E. MgrB alterations mediate colistin resistance in Klebsiella pneumoniae isolates from Iran. Frontiers in Microbiology. 2017;8:1-8
21.Gao W, Chua K, Davies JK, Newton HJ, Seemann T, Harrison PF, et al. Two novel point mutations in clinical Staphylococcus aureus reduce linezolid susceptibility and switch on the stringent response to promote persistent infection. PLOS Pathogens. 2010;6:e1000944
22.Kojima S, Nikaido H. Permeation rates of penicillins indicate that Escherichia coli porins function principally as nonspecific channels. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:2629-2634
23.Tran QT, Williams S, Farid R, Erdemli G, Pearlstein R. The translocation kinetics of antibiotics through porin OmpC: Insights from structure-based solvation mapping using WaterMap. Proteins. 2013;81:291-299
24.Djahmi N, Dunyach-Remy C, Pantel A, Dekhil M, Sotto A, Lavigne J-P. Epidemiology of carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Mediterranean countries. BioMed Research International. 2014;2014:11
25.Li B, Zhao Y, Liu C, Chen Z, Zhou D. Molecular pathogenesis of Klebsiella pneumoniae. Future Microbiology. 2014;9:1071-1081
26.Hu R-M, Liao S-T, Huang C-C, Huang Y-W, Yang T-C. An inducible Fusaric acid tripartite efflux pump contributes to the fusaric acid resistance in Stenotrophomonas maltophilia. PLoS One. 2012;7:e51053
27.Bialek-Davenet S, Lavigne J-P, Guyot K, Mayer N, Tournebize R, Brisse S, et al. Differential contribution of AcrAB and OqxAB efflux pumps to multidrug resistance and virulence in Klebsiella pneumoniae. Journal of Antimicrobial Chemotherapy. 2015;70:81-88
28.Naas T, Oueslati S, Bonnin RA, Dabos ML, Zavala A, Dortet L, et al. Beta-lactamase database (BLDB)—Structure and function. Journal of Enzyme Inhibition and Medicinal Chemistry. 2017;32:917-919
29.Livermore DM. Defining an extended-spectrum beta-lactamase. Clinical Microbiology and Infection: The Official Publication of the European Society of Clinical Microbiology and Infectious Diseases. 2008;14(Suppl 1):3-10
30.Bush K, Jacoby GA. Updated functional classification of beta-lactamases. Antimicrobial agents and chemotherapy. 2010;54:969-976
31.Öztürk H, Ozkirimli E, Özgür A. Classification of beta-lactamases and penicillin binding proteins using ligand-centric network models. PLoS One. 2015;10:e0117874
32.Navon-Venezia S, Kondratyeva K, Carattoli A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiology Reviews. 2017;41:252-275
33.Gupta A, Ampofo K, Rubenstein D, Saiman L. Extended spectrum beta lactamase-producing Klebsiella pneumoniae infections: A review of the literature. Journal of Perinatology: Official Journal of the California Perinatal Association. 2003;23:439-443
34.Zhang J, Zhou K, Zheng B, Zhao L, Shen P, Ji J, et al. High prevalence of ESBL-producing Klebsiella pneumoniae causing community-onset infections in China. Frontiers in Microbiology. 2016;7:1830
35.Toubiana J, Timsit S, Ferroni A, Grasseau M, Nassif X, Lortholary O, et al. Community-onset extended-spectrum β-lactamase-producing Enterobacteriaceae invasive infections in children in a University Hospital in France. Medicine. 2016;95:e3163
36.Hawkey PM. Prevalence and clonality of extended-spectrum-beta-lactamases in Asia. Clinical Microbiology and Infection: The official Publication of the European Society of Clinical Microbiology and Infectious Diseases. 2008;14:159-165
37.Zamorano L, Miro E, Juan C, Gomez L, Bou G, Gonzalez-Lopez JJ, et al. Mobile genetic elements related to the diffusion of plasmid-mediated AmpC beta-lactamases or carbapenemases from Enterobacteriaceae: Findings from a multicentre study in Spain. Antimicrobial Agents and Chemotherapy. 2015;59:5260-5266
38.Naas T, Cuzon G, Villegas MV, Lartigue MF, Quinn JP, Nordmann P. Genetic structures at the origin of acquisition of the beta-lactamase blaKPC gene. Antimicrobial Agents and Chemotherapy. 2008;52:1257-1263
39.Lee CR, Lee JH, Park KS, Kim YB, Jeong BC, Lee SH. Global dissemination of carbapenemase-producing Klebsiella pneumoniae: Epidemiology, genetic context, treatment options, and detection methods. Frontiers in Microbiology. 2016;7:895
40.Qamar MU, Saleem S, Toleman MA, Saqalein M, Waseem M, Nisar MA, et al. In vitro and in vivo activity of Manuka honey against NDM-1-producing Klebsiella pneumoniae ST11. Future Microbiology. 2018;13:13-26
41.Qamar MU, Walsh TR, Toleman MA, Saleem S, Jahan S. First identification of clinical isolate of a novel “NDM-4” producing Escherichia coli ST405 from urine sample in Pakistan. Brazilian Journal of Microbiology [Publication of the Brazilian Society for Microbiology]. 2018;49:949-950
42.Toleman MA, Spencer J, Jones L, Walsh TR. blaNDM-1 is a chimera likely constructed in Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy. 2012;56:2773-2776
43.Jeong SH, Lee KM, Lee J, Bae IK, Kim JS, Kim HS, et al. Clonal and horizontal spread of the blaOXA-232 gene among Enterobacteriaceae in a Korean hospital. Diagnostic Microbiology and Infectious Disease. 2015;82:70-72
44.Kilic A, Baysallar M. The first Klebsiella pneumoniae isolate co-producing OXA-48 and NDM-1 in Turkey. Annals of Laboratory Medicine. 2015;35:382-383
Written By
Muhammad Usman Qamar, Muhammad Hidayat Rasool, Shah Jahan,
Muhammad Shafique and Bilal Aslam
Reviewed: October 29th, 2018Published: April 3rd, 2019